「脂質」とは一般的には水に溶けにくく、油に溶けやすい物質のことを指します。脂質には様々な機能がありますが、そのうち重要なものの一つがシグナル分子としての機能です。例えば脂質の一種である「リン脂質」は生体膜を構成する成分の一つですが、細胞が様々な刺激を受けると「膜から切り離され」、細胞内や細胞外に遊離し、情報を伝える物質として機能します。この「膜から切り離す」すなわち「活性化」の過程を担っているのがリン脂質を加水分解する「ホスホリパーゼ」と呼ばれる一群の酵素です。
これまで動物ではホスホリパーゼ群が関わる脂質シグナル分子の産生経路およびその機能が明らかになっています。例えば細胞がホルモンやサイトカインなどの刺激を受けるとホスホリパーゼが活性化し、その下流でプロスタグランジン・ロイコトリエンなど、いわゆる「エイコサノイド」と呼ばれる脂質メッセンジャーが産生され、それらが受容体を介して血管収縮、血小板凝集、炎症など多様な応答を引き起こすことがわかっています。
一方、微生物ではこうした「脂質シグナル」の存在はあまりよくわかっていません。麹菌のゲノムには動物と類似した様々なホスホリパーゼ群が見出されていますが、それらがどのような働きをしているのかは謎に包まれています。当研究室では、麹菌を含めた微生物において「脂質シグナル」伝達経路がどのような役割を果たしているかを明らかにし、その知見を育種や物質生産などに応用することを目的に、麹菌の持つホスホリパーゼ群の解析を行ってきました。その過程で、麹菌の持つホスホリパーゼが動物のものとはかなり異なり、ユニークな性質・機能を持つことがわかってきました。
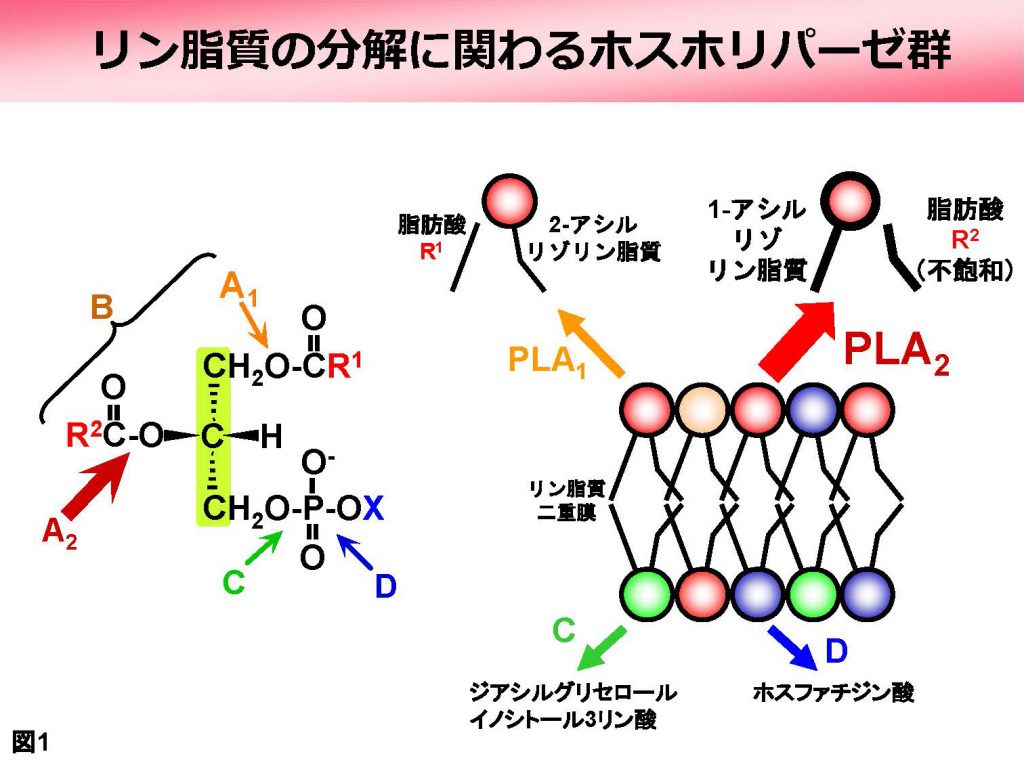
「ホスホリパーゼ (phospholipase)」はその名の通りリン脂質を加水分解する酵素ですが、分解様式によって様々なタイプが知られています(図1左)。このうち「ホスホリパーゼA2(PLA2と略)」はリン脂質のグリセロール骨格(緑の部分)に結合した2つの脂肪酸(R1とR2)のうち、2番目の結合だけを特異的に加水分解します。その結果、R2の脂肪酸と、R1が残った部分(これを1−アシル型のリゾリン脂質と言います)ができることになります(図1右上)。
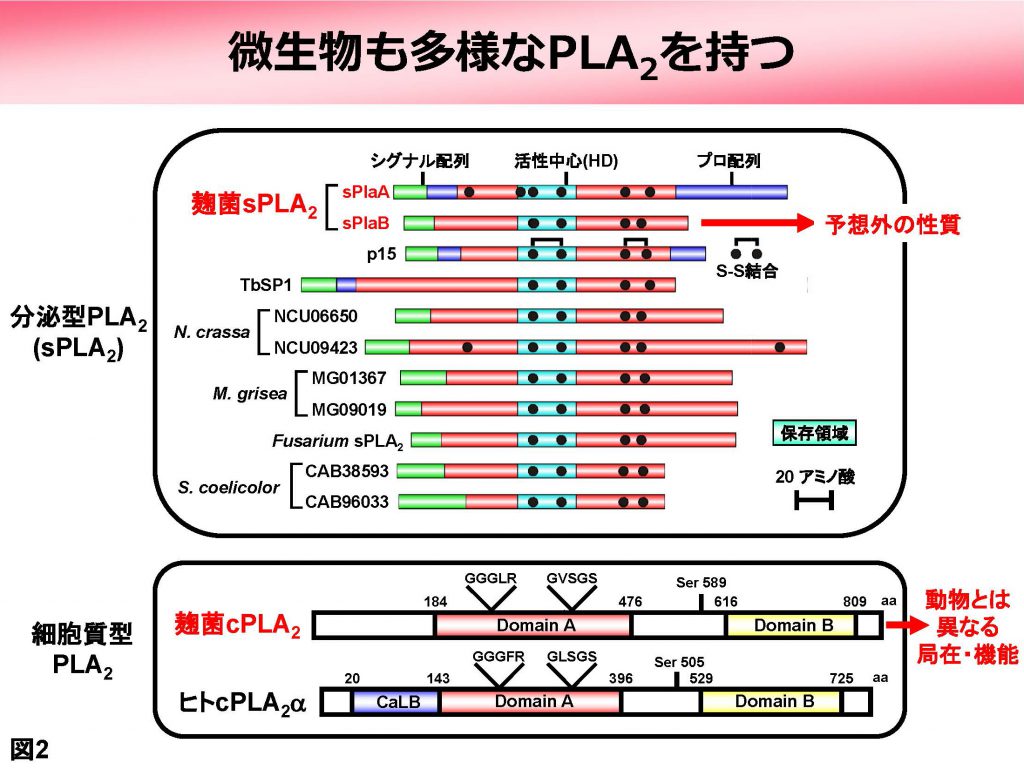
PLA2はさらにいくつかのサブタイプに分類されます。それらは、細胞外に分泌される「分泌型 (secretory PLA2 (sPLA2))」、細胞質にある「細胞質型 (cytosolic PLA2 (cPLA2))」、そして「カルシウム非依存型 (calcium-independent PLA2 (iPLA2))」などです。これらは動物では古くから知られていましたが、我々は麹菌がsPLA2とcPLA2のオルソログを持つことを見出しました(図2)。それらがどのような働きを持つかを解析したところ、以下に述べるように、動物のオルソログとはかなり異なる性質・機能を持つことがわかってきました(図3)。
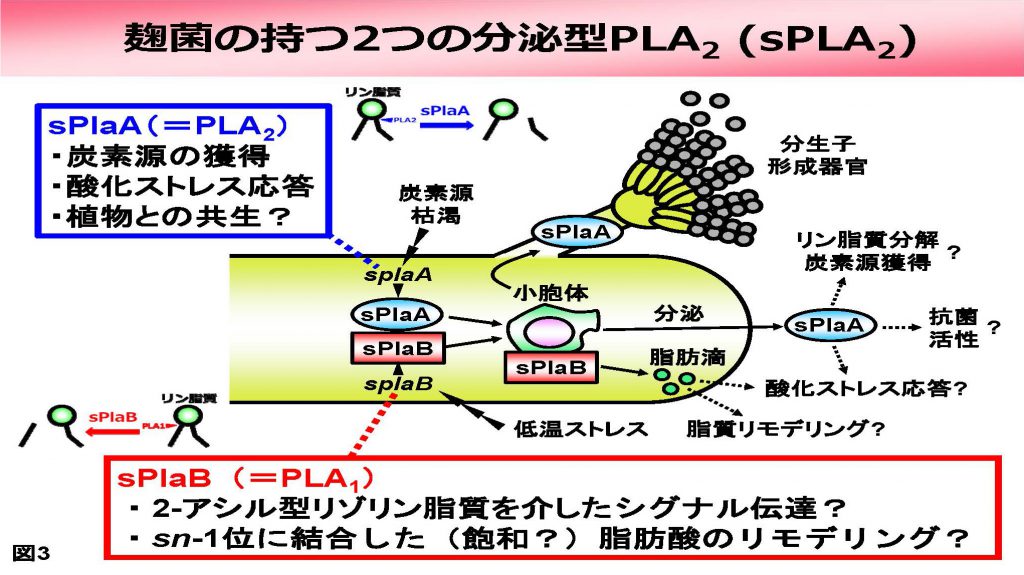
まず分泌型であるsPLA2ですが、麹菌はsPlaA、sPlaBと命名した2つのタンパク質を持っています。両者はアミノ酸配列上は互いによく似ていますが、その局在は異なり、sPlaAが細胞外に分泌され、また分生子形成器官にも認められるのに対し、sPlaBは細胞内の小胞体あるいは脂肪滴と考えられるオルガネラに局在していました。
また両者は活性の至適Ca2+濃度やpH、発現が誘導される条件なども異なっており、重複した遺伝子ではなく、独立した機能を果たすものと考えられました。また最近、sPlaBがPLA2ではなく、実は1番目の結合を切断するホスホリパーゼA1(PLA1)であるという、実に意外な結果が得られました(図4)。
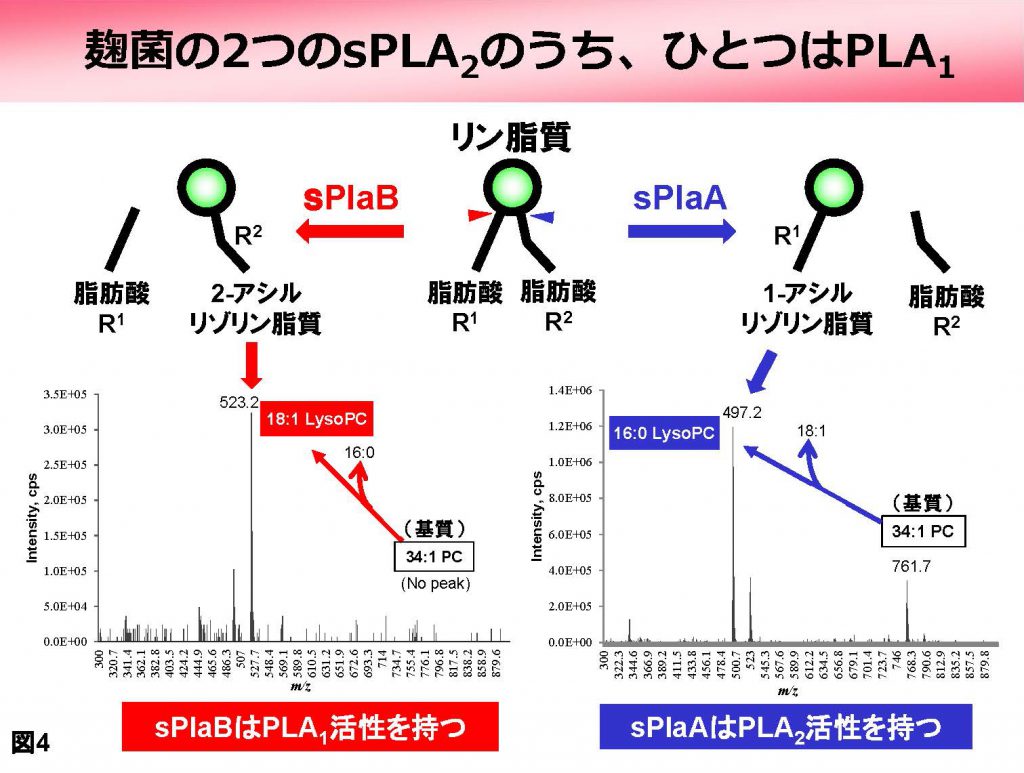
さらに調べると、別のカビが持つ「sPLA2」の中にも実はPLA1が存在することがわかりました。これまで動物由来を含め数十のsPLA2が単離・解析されていますが、いずれも厳密に2番目の結合だけを切断すると言われており、PLA1としての報告は皆無です。一般にリン脂質のR1は飽和脂肪酸、R2は不飽和脂肪酸であるることから、sPlaBの作用によってR1由来の飽和脂肪酸と2番目の位置に不飽和脂肪酸を持つ特殊な2−アシル型のリゾリン脂質が産生されることになりますが、それらの役割は全く不明です。
一方、細胞質型であるcPLA2のオルソログですが、これは麹菌ゲノムからは1個見つかり、AoPlaAと命名しました。動物ではcPLA2は通常は細胞質にいますが、刺激を受けると細胞内のオルガネラに移行し、その膜を切断することが知られています(図5)。
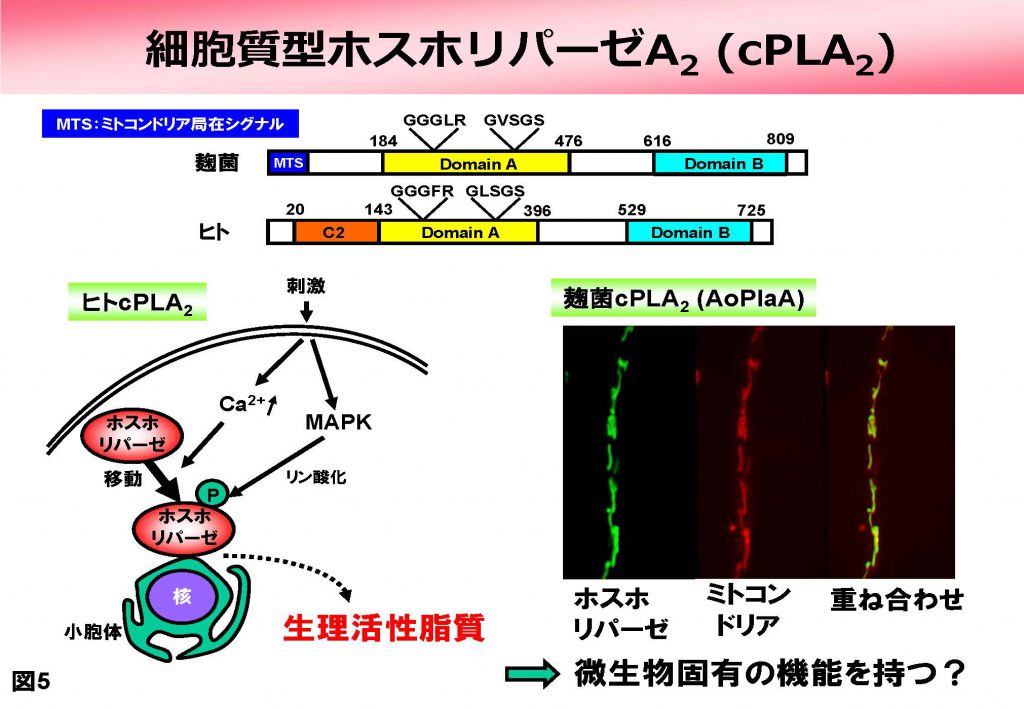
ところが、麹菌オルソログであるAoPlaAの場合、N末端にミトコンドリア局在シグナル(MTS)を有しており、ミトコンドリア内膜と外膜の間の膜間スペースに局在することがわかりました。また、AoPlaAは様々なリン脂質のうちホスファチジルエタノールアミンとカルジオリピンのみを特異的に分解する、ユニークな基質特異性を持つことがわかりました。これらの知見は動物cPLA2で得られているものとは全く異なっており、AoPlaAが動物cPLA2とは異なる固有の機能を果たすことが強く示唆されました。
以上のように、麹菌の持つPLA2は動物のオルソログとアミノ酸配列上はある程度似通っているものの、その性質・機能はかなり異なることがわかってきました。これらの酵素が、その産物である脂肪酸やリゾリン脂質の産生を通じてどのような「脂質シグナル」を発していて、それが麹菌(や他の糸状菌)においてどのような役割を果たしているのかを解明したいと考えています。