Gタンパク質共役型受容体(G protein-coupled receptor; GPCR)は細胞膜上に存在する最大の受容体ファミリーであり、細胞膜を7回貫通する特徴的な構造を持ちます。GPCRは特異的なリガンドや光といった細胞外からの刺激を感知し、細胞内領域に結合したGタンパク質をGDP型からGTP型に変換して活性化することで細胞質内へシグナルを伝達します。
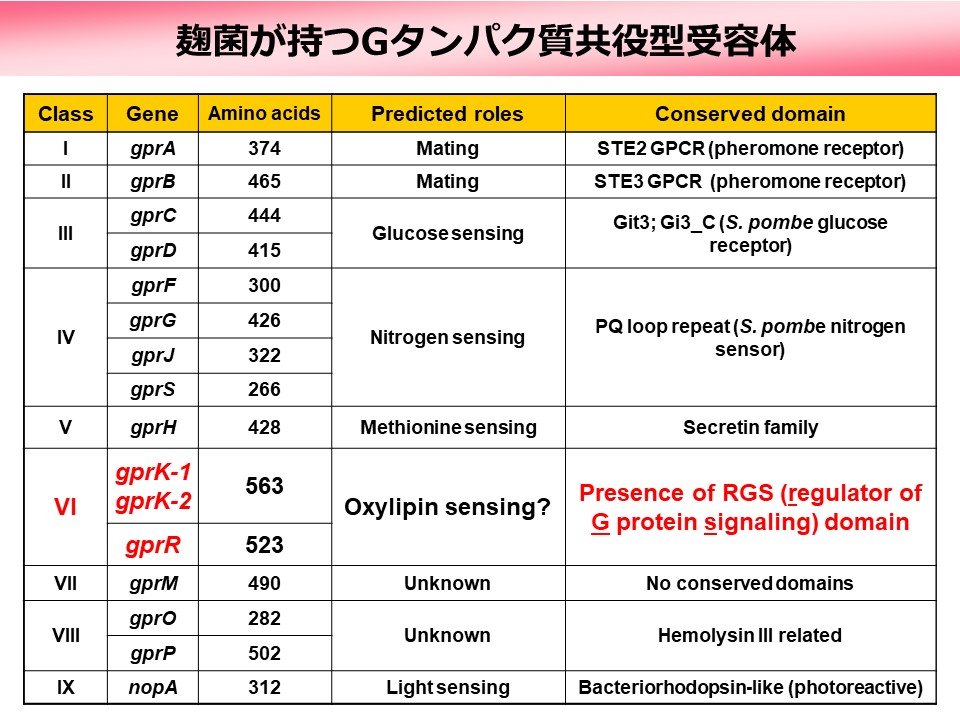
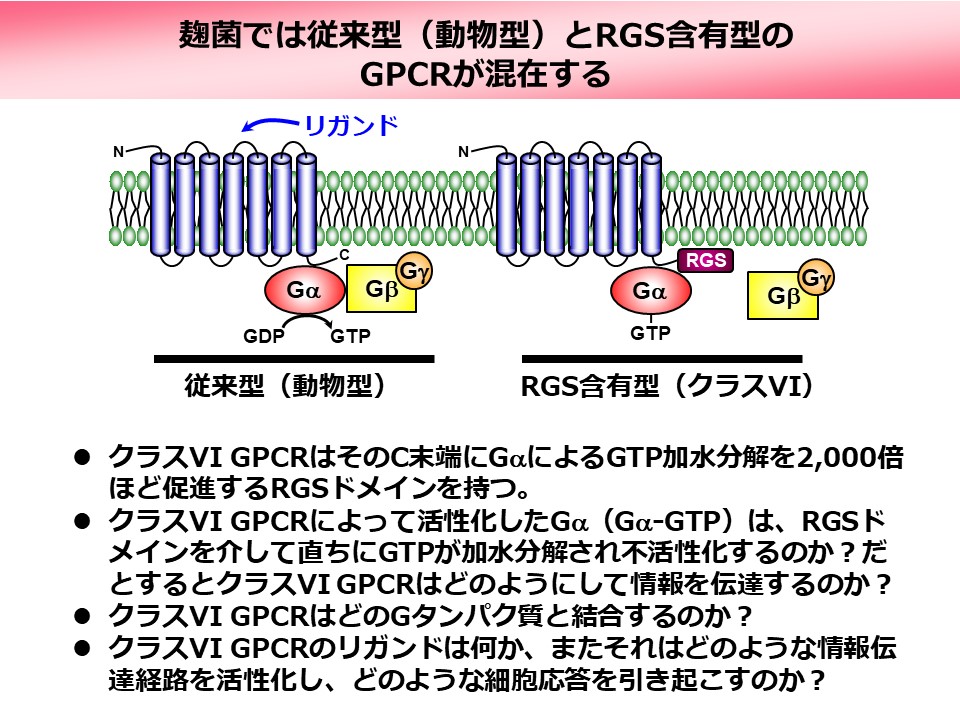
糸状菌である麹菌Aspergillus oryzaeは16種類のGPCRを持ち(上図)、機能や相同性によってIからIXの9つのグループに分類されていますが、リガンドや細胞内情報伝達経路などの詳細には不明な点が多く残されています。我々はそれらのうちクラスVIに属する3つのGPCR(GprK-1、GprK-2、GprR)に着目しています。その理由は、これらのGPCRがC末端にRGS(Regulator of G protein Signaling)ドメインを持つ特殊なGPCRであるからです。RGSドメインはGタンパク質の加水分解を強く促進する機能を持つことから、クラスVIのGPCRはGタンパク質の活性化(GDP型からGTP型への変換)と不活性化(GTP型からGDP型への変換)という正反対の反応を司る領域を一つの分子内に持つことになります(下図)。このような例は動物のGPCRでは知られておらず、その制御や機能がどのようなものであるか興味が持たれます。最近我々は麹菌のクラスVI GPCRが菌核と呼ばれる特殊な構造体の形成に関わることを見出しました。それがどのようなシグナル伝達経路を介したものであるか解明を進めています。